THE BONUS ROUND PRESENTS
Dickie's Personal HIV TheoryThe FULL MONTY - Chapter 3
by Dick Remley, every doctor's
nightmare:
A PATIENT WITH AN INFORMED OPINION
|
CHAPTER THREE: THE "TECH" STUFF HIV Disease and The THEORY- In
A Nutshell
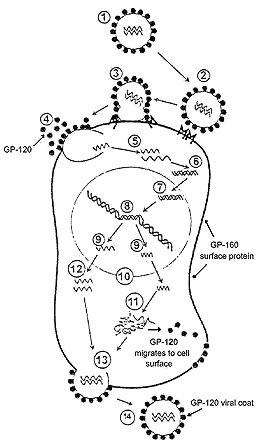
Okay, now we're going to get into the mechanics of this. This chapter will deal with the technical details of The THEORY and its description of the process by which HIV causes clinical disease. This is not intended to be a textbook on microbiology. I will still attempt to simplify things by leaving out extraneous issues that may be true in detail, but don't really impact this discussion. (Such as how viruses other than HIV might replicate or cause disease.) This is also my weak suit. Not being trained in microbiology or immunology, I am dealing with an area in which I admit my knowledge is incomplete. By way of fairness, however: EVERYBODY'S knowledge of microbiology and immunology is incomplete. New discoveries are made daily, and the field can change literally overnight. I present The THEORY as a means of grasping the complex issues of HIV infection. It is meant to be a reasonably predictive model. Much like the idea of the structure of an atom being like a tiny solar system: untrue in detail, but workable on many practical levels. With that in mind, let us proceed onward. First, we need some background information: IMITATION OF LIFE ? Viruses have traditionally been thought of as very simple things. So simple, in fact, that there has been past debate as to whether viruses qualify as life-forms at all. Often, they are thought of as just a bag of chemicals randomly floating around until they bump into a host cell and break open. It's not quite that easy. What seems to fuel the debate about whether viruses are living things, or whether they merely MIMIC living things, is the issue of reproduction. Viruses cannot reproduce on their own: they lack the necessary equipment. They "borrow" that equipment from the cells of the host they infect. Viruses merge with those cells in one way or another in order to produce offspring. In some ways, that makes viruses a lot like men: not really self-sufficient, they just roam around looking for a chance to procreate. Ask any woman. Actually, there ARE similarities between viral reproduction and sexual reproduction. In some ways, the virus is like a sperm cell, and the host cell is like an egg. But we're not going to debate these similarities. They are somewhat superficial. Like men. Ask any woman. Okay, okay...I know: what's with all these "men are like a virus" jokes? Well, in an effort not to get too sidetracked by the issue of whether or not viruses are living things, I'm going to take the position that viruses are no less alive than men, and put that whole debate aside. I'm going to refer to viruses as if that issue has been settled, and we've decided viruses are living things. It's mostly definitional, and irrelevant to the other issues we're going to discuss here. I just wanted to get it out of the way. Later on, I'm going to give men a break by claiming that they really DO do more than just reproduce. I'm going to claim the same thing about the HIV virus. PROTEIN SYNTHESIS: TEENAGERS IN LOVE I think most of you will probably recall from high school biology that, inside the nucleus of each cell there is a double-stranded molecule that determines our genetic make-up. It is called DNA, and it is twisted into a double-helix formation. I'm not going to go into the details of how genes are composed of DNA, and how DNA is composed of nitrogen-based nucleosides attached to sugar molecules and phosphate groups. I tried doing that in a rough draft of this work, and, trust me, it was boring as all heck. If you want to know the details of all that stuff, E-mail me and I'll tell you about it. All I want you to remember right now is that, when a cell divides, the two strands of DNA separate, and the molecule "unzips" lengthwise down the middle. This exposes the nucleoside components of the molecule, which then act as templates for the formation of new copies of the DNA. But this is not the only thing DNA does. Many times, the DNA will not completely separate from end to end; but will only partly unzip starting and ending somewhere in the middle of the molecule. When it does this, protein synthesis begins. In protein synthesis, the exposed nucleosides in the middle of the DNA molecule serve as templates for the formation of RNA rather than DNA. The nucleosides of RNA then serve in groups of three (called codons) as receptor sites for amino acids which are carried to those sites by another, slightly different form of RNA. Amino acids can behave kind of like teenagers in love: when they get close to one another, they want to "hold hands". But there are atoms on each side of the amino acid molecules that prevent that. So, along comes a special enzyme that removes those atoms. The atoms that are removed are hydrogen and oxygen, in a ratio of two-to-one. Once the hydrogen and oxygen are out of the way, the amino acids link up by forming a special attachment to one another. This attachment is called a "peptide bond". After the amino acids along the RNA molecule are all strung together like pearls on a necklace, they are released in this string-like formation. But wait! What about the hydrogen and oxygen atoms the enzymes freed from the amino acids? Well, they come together and form water molecules (H2O). Here's where things get interesting. (You mean they're FINALLY going to get interesting?): Water molecules are polar. No, that doesn't mean it's ice water - "polar" in another sense. Water molecules are not symmetrical. The hydrogen atoms tend to be grouped off to one side of the molecule, leaving the other side to be mostly oxygen. Hydrogen and oxygen have different electromagnetic charges, and this makes the water molecule polar like a magnet: one end is positively charged, and the other end is negatively charged. Amino acids also have an electromagnetic structure, but the amino acids are not all alike. There are 20 amino acids, and when they line up in this peptide chain we've described, some of the amino acid structures are attracted to the water that is formed, and some are repelled by it. There is a kind of push-pull, attract-and-repel activity that occurs at this point. The amino acid chain is forced to kink, coil and fold until it finds a stable shape. This "dance" between the amino acids and the surrounding water limits the number of choices of shapes the molecule has. In fact, it limits it to about one choice, dependent on its amino acid composition. Even though these molecules, when seen with electron microscopes and x-ray crystallography appear to have about as much shape as a ball of lint, that shape is consistent with each molecule's amino acid sequence. These "folded-up" strands of amino acids are called "proteins". Their individual shapes largely determine the uniqueness of their functions. Besides water, most of the parts of living things - including you and HIV - are made of proteins. Proteins are often called something else, depending on their form and function. For example, some proteins are called "enzymes". Enzymes are proteins that facilitate or speed up chemical reactions in the body. Think of them like the flame on your stovetop burner. You could dump a whole bag of unpopped popcorn into a pot with some oil, put it on the burner, and wait for the temperature of the air in the room to get high enough to pop the corn; but you're gonna wait a really long time before that corn pops. And then, if it ever DOES pop, you'd better run like heck, because it probably means the kitchen is on fire! However, if you turn the burner's flame on, the corn pops quickly as soon as it reaches the right temperature range; and you remain relatively unscathed. Not a perfect analogy... but you get the point. Many chemical reactions in your body would require too much heat to occur safely within your cells. Your body would burn up if enzymes didn't initiate and contain these processes, making them faster and easier. You should also know that the amino acids in a protein can be linked to other types of molecules - like fats, sugars, metals, etc. Scientists sometimes categorize proteins into groups named for these other molecules. For example, a protein in which the amino acids are linked to sugar molecules can be referred to as a "glycoprotein". So, why am I telling you all this? Well, I thought it might be helpful to your ability to understand the rest of The TECH Stuff... VIRAL REPRODUCTION There are different kinds of viruses and they reproduce somewhat differently, depending on their type. We are only going to discuss HIV here. When HIV successfully infects a cell, it does so by using protein structures on its surface to attach to protein structures on the host cell's surface. Once attached, the virus "fuses" with the host cell and injects an inner capsule-like structure (called the "capsid") into the interior of that cell. The outer protein coat of the HIV is discarded by the virus and left behind. That outer coat may then break up and float off into the bloodstream. Now, if those proteins are floating around in the bloodstream, how is the body to differentiate between them and a fully formed copy of the virus? (Fully formed viral structures are referred to as "virions" so as to distinguish them from the various individual parts into which the virus breaks down.) HIV MUST break down in order to incorporate itself into the cell. This sometimes makes clarity of discussion difficult in regard to HIV, because at some point the virus has scattered bits and pieces of itself around and deposited them in various places. It can eventually become difficult to define just where the virus is, and just exactly what it consists of. Might this not confuse the body's immune system? What should the body do? Where should it attack? Should it attack while the virus is still inside its protein coat - or wait until the virus has fully disassembled itself? Why not leave the part of the virus inside the cell for the cell itself to deal with (by apoptosis, for example), and hone in on the outer protein coat? The immune system attacks in as many ways as it can. But, if much of the viral protein in the bloodstream is actually made up of discarded shell left behind by the virus, wouldn't the body's immune system be over-responding? Could this not hyperactivate the immune system needlessly? This is part of what I meant earlier when I said the body might be responding to a "false signal" sent by the virus. Host-produced antibodies might be expending a lot of wasted energy cleaning up what is really just so much discarded junk. If that "junk" were to lodge in tissues, it might be possible that the body would begin to attack those tissues needlessly. Unable to distinguish between the fully assembled virions and the useless junk, the body would respond by kicking into "hyperdrive" in order to cover all bases. This might be one way in which HIV causes autoimmune disease and allergy-like symptoms. Meanwhile, as the body is busy rounding up all this "junk", the really effective part of the virus sinks quietly into the cell. Once inside the cell, the capsid containing the parts of the virus essential to replication migrates toward the cell's nucleus where the virus undertakes the process of incorporating itself into the DNA - the host's genetic material. In so doing, the virus once again removes its outer surface - this time the capsid shell. The capsid's contents (two strands of RNA and a few enzymes) emerge to integrate themselves into the host's DNA structure. HIV accomplishes this task with the help of a couple of the enzymes it carries along with it - one called "Reverse Transcriptase" and one called "Integrase". (Whenever you see the suffix "ase" at the end of a word, it generally means that word is describing an enzyme.) The reverse transcriptase "reads" the RNA strands as a template to create a DNA image of itself, which is then integrated into the host's own DNA structure. Successfully integrated, the original distinguishable virus has now all but disappeared. At this point, HIV has become a "pro-virus" - a sort of "machine" that may lie dormant, or may use the host cell's ability to replicate in order to crank out copies of the proteins the virus uses to assemble itself. Once again referring back to previous parts of this work: this development of provirus is what I meant when I referred to the "successful" infection of a cell or to "successfully infected" cells. Another enzyme the virus uses is called "Protease". Protease facilitates a process by which a cellular protein called GP-160 is cut up into GP-120 and GP-41. "GP" just stands for "glycoprotein", and the number following it tells you by weight, how many amino acid - sugar structures are in its composition. You may notice that the numbers don't add up: GP-160 minus GP-120 would equal GP-40, not GP-41. This is because some dummy didn't round things off correctly and GP-160 is REALLY GP-161. [ Okay, now before I start getting hate mail from that dummy and all of his or her supporters, let me acknowledge that this was really due to a problem with the sensitivity level of the equipment used to measure the components, rather than to a mistake by the observer. So don't go judging them as harshly as I just did. Sometimes I have problems with the sensitivity of MY equipment, too, you know. ] As far as I can tell, scientists let the GP-160 misnomer stand because... well, because... well, because they seem to get a kick out of knowing something you don't know, and leaving things misnamed allows them to maintain this feeling of superiority. [Whoops! There goes the sensitivity level of my equipment again!] Anyhow, GP-160 is a part of the surface of the cell, and migrates to the exterior of the cell where it belongs. When GP-160 is cut down to GP-120, the GP-120 also migrates to the cell surface. The difference is that, while GP-160 belongs there, GP-120 does not. In fact, GP-120 really has no business being on the cell's surface at all... but for one little detail: GP-120 is the protein that makes up the outer coat of the HIV virion. At the end of its replication cycle, HIV collects together its necessary components and, along with a newly formed capsid, migrates to any portion of the cell surface that is composed of GP-120. The virus then uses the GP-120 as an exiting point from the cell. It breaks out of the cell by wrapping itself in the GP-120. This process is often described as "budding". The newly born virion then floats off into the bloodstream, shrouded in its new GP-120 coat. The following graphic illustrates the process in a simplified form:
How, then, could HIV infect new cells? Well, remember how we discussed that HIV must use its outer protein structure to attach to a cell's surface? And remember how we discussed that the outer surface of HIV is formed out of GP-120? And remember, too, that HIV infected CELLS display GP-120 on their surfaces? Well, what if the GP-120 on the cell surface allowed successfully infected CELLS to attach to other cells and infect them directly without HIV ever having to emerge into the bloodstream? In this way, cells could clump together and HIV could move from cell nucleus to cell nucleus rather than from cell surface to cell surface, avoiding the bloodstream altogether. The next issue, of course, is obvious: What's to prevent the body from identifying the GP-120 on an infected cell's surface, and attacking the cell directly? There are some complex explanations for this - one of which involves a gene called Vpr - but, you know me and my distaste for complex explanations. Instead, I'm going to offer a very simple one: What if the cell simply leaves the bloodstream and migrates - or marginates - into tissues where it cannot be reached very well by the circulating antibodies? Getting "out of the line of fire", so to speak. The result of all this would be that the body expends a lot of immunological energy cleaning up "junk" GP-120 left behind by virions attempting to infect cells directly from the bloodstream, while the successfully infected cells slip away into the surrounding tissues. In oversimplified terms: the virus throws up a smoke screen while infected cells slip out the back door. How efficient would this process be? Not very. In order to pass on the virus, the infected cells imbedded in the tissues would have to rely on drawing other cells to them. Meanwhile, the GP-120 on the infected cell's surface might make it vulnerable to other immunological defenses. Only a certain small percentage of infected cells would find a safe hiding place. It would likely take a long time before the virally infected cells caused immunological collapse. Like, oh, say... about eleven years or so. Meanwhile, the host would often not feel well, and might begin developing autoimmune and allergy-like symptoms. Sounds like HIV disease to me. Of course, the next problem is to account for the body's failure to replace the infected T-cells. You COULD say that this effect is simply the result of a slow process. But it also seems possible to me that the cells might need to send out a signal for replacement, and that HIV prevents this. Of course, THEN you'd have to explain HOW HIV prevents that. Well, I'm going to take a shot at that explanation. (You didn't think I could resist THAT did you?) OUT ON A LIMB Remember how I promised to vindicate men a bit by taking them out from under the "all they do is wander around looking for a chance to reproduce" label I've stuck on them? Well, this is it: I seem to remember from back in my high school biology days that the criteria for determining whether or not something is a living thing consists of several factors, not just whether the thing can reproduce. In fact, from an evolutionary point of view, living things have to do two critical things in order to survive as a species: 1.) They must produce offspring; and 2.) They must be able to defend themselves so they can live long enough to reproduce. They can defend themselves either by fighting or running away. Men do that. Men have the urge to merge and the urge to fight. Then they often run away. They are constantly causing trouble and then trying to making things all right again by having sex. Fighting and making up... that's how guys organize their relationships. See? Guys ARE good for something other than sex: they're good for fighting with, too. Ask any woman. Sorry, guys: I guess that wasn't quite as complimentary as we all had hoped. The HIV virus behaves similarly: it attacks the cell, then merges with it. HIV is incompletely equipped for either reproduction or self-defense, so it must borrow both functions from the cell's already existing apparatus. But just what PART of the cell's defense apparatus could it use? This is where we're going to get into cytokines a bit. I've mentioned them before, and we're now going to take a brief but closer look at them: Cytokines are tiny bits of protein that connect to receptor sites on cells and that can stimulate some portion of the immune response. Cytokines like Tumor Necrosis Factor (TNF) start the inflammatory cascade. Blood vessels dilate, and the walls of those blood vessels become more permeable, allowing white cells to travel across this barrier more freely. Cytokines such as IL-2 cause the proliferation of certain white cells (including CD-4's) so there are more of them available to attack infection. However, I don't think IL-2 causes the creation of new T-cells so much as it causes existing T-cells to divide. The reason for me thinking that is the phenomenon of viral load increase: When IL-2 is used to cause T-cell proliferation, it also causes HIV viral load to go up. This is what you would expect to see if cells carrying the HIV provirus were stimulated to divide. IL-2 is a useful tool, but only if you can suppress viral replication at the same time you administer IL-2... otherwise, it's kind of a wash. Other cytokines assist in the inflammatory process as well. Cytokines offer us a model of a mechanism for activation of the immune system, and for cellular replication. To create brand new cells, a cytokine would have to stimulate cellular production in the bone marrow, which is ultimately where all blood cells originate. It is possible to think of cytokines as being part of the cell's defense system: the part that sends for help. Now, if the cytokines are inducing and regulating inflammatory responses, they can't just be present in your bloodstream in significant numbers all the time. If that were the case, you'd walk around inflamed all the time. The cytokines must be coming from somewhere. Obviously, they are coming from somewhere in the cells. This, then, means that the cells must produce cytokines. But are the cytokines being produced and stored inside the cell, or are they not produced until infection occurs? The following is sheer guesswork on my part: Logically, it makes more sense for cells to produce and STORE cytokines so they can be released as rapidly as possible when a cell is attacked by an outside invader. Where would the cell store these cytokines? Again, the logical answer would be: "in the location where they would be most readily available". In other words, they must be at or near the cell surface, so that any initial breach in that surface would immediately induce inflammation. Well, what if they are ON the cell's surface? What if they are strung together so they function as ONE kind of protein while the cell is intact, and like a DIFFERENT kind of protein when the cell's surface is disrupted? Breaching the cell surface would break the amino acid chain of the protein string, disrupt the electromagnetic balance of the structure, and cause the protein and any fragments of it to re-fold, giving them a new shape. Since the shape of a protein largely determines its function, these fragmented proteins could take on a new role. Perhaps a cytokine-like role. And what if this theoretical string of cytokines forms a surface protein commonly known as GP-160? This would make the amino acid sequences in GP-160 an essential part of the cell's ability to induce inflammation and stimulate cell growth. But if GP-160 were cut down into a smaller chain of amino acids - say, GP-120 - wouldn't some of that amino acid sequence now be missing? And what if the missing sequence was part of a cytokine that stimulates new T-cell production? Then the cell would become deficient in it's ability to signal T-cell production. It's just a theory. And certainly not essential to The THEORY as a whole. But what if it's right? The implication would be that there might be a whole other approach to HIV treatment: using anti-inflammatory agents to cut down on autoimmune symptoms and slow the syncytial process, one could then administer doses of the proper T-cell producing cytokine and restore near-normal immunity without ever having to directly attack the HIV virus at all. In other words, you'd assist in the creation of a symbiotic relationship, and allow the virus and the host to co-exist. Live and let live. What a concept! Of course, you couldn't constantly immunosuppress the host unless you had an effective suppressant that worked only on the overproduced inflammation, as opposed to ALL the body's inflammatory processes. Using current commonly available immunosuppressants, you'd have to go on and off them every few weeks to allow the body to fight infections other than HIV. It would be interesting to compare the amino acid sequence of GP-160 to known cytokines to determine if the process I described above might be possible. And it would be at least equally as interesting to compare the amino acid sequence of GP-160 to GP-120 to see what part is missing or chopped up. Just a thought. Okay, we are now at the point where we can once again summarize The THEORY, but in detailed terms, and with the addition of some predictable consequences. The THEORY When HIV enters the bloodstream, it may do so as free-floating virions, or it may gain entry as a provirus inside an infected cell. This implies that co-infection with a disease that draws white cells to the site of transmission (usually the genital area) would increase the risk of transmission significantly. In other words: you'd be more likely to spread HIV if you also have a venereal disease. Other non-venereal inflammations in the pelvic area would also increase the risk for both parties involved. It takes a little while for the body to produce an antibody to the HIV virions. During this stage, HIV would more easily move from cell-surface to cell-surface through the bloodstream. Also, provirally infected host cells that have expressed HIV-created proteins such as GP-120 on their surfaces would have a better opportunity to become syncytial and migrate out of the bloodstream. One of the major factors in the body's defense against the virus at this point would be apoptosis. Because of this, T-cell counts would fall dramatically in the time just following initial infection. Once the antibody to HIV appears, HIV's chances of survival in the bloodstream would drop significantly. It might occasionally infect a cell through the circulatory system and then go on to create a provirus, but the odds would be against it. As the immune system gears up, the infection of new cells would be severely curtailed. T-cell decline would slow down at this point. In fact, the number of T-cells would perhaps even RISE for a while after the body begins antibody production; but T-cell counts would never rise completely back to normal. Some provirally infected cells would already have become syncytial and migrated or marginated out of the bloodstream into the tissues of the lymphatic system, the liver, the spleen and into the brain-blood barrier, as well as to other sites. These cells would have a slight edge in survival, but it would take years for such a small survival advantage to greatly impact the host. Among other things, the process described above implies that it would be unlikely for a person already infected with HIV to become re-infected by someone else. Free-floating fragments of GP-120 would continue to hyperactivate the immune system. This, coupled with inflammation induced by the marginated or sequestered proviral cells, would frequently induce symptoms of allergy or autoimmune disease. In addition to other things, this might cause Polyglandular Deficiency Syndrome by altering adrenal function. Because of this, an HIV infected individual might feel very ill for years, despite continually demonstrating lowered, but adequate T-cell counts. Eventually, the syncytial nature of the marginated and sequestered proviral cells would allow them to absorb other white cells drawn to them by inflammation. This absorption of cells would prevent them from alerting the body that those cells need to be replaced, and cell counts would begin to decline more rapidly. An HIV-induced deficiency in proteins necessary to send this "alert signal" might slow the process of T-cell replacement even more. With the decline in T-cell counts accelerating, the host would lose his ability to fight off other illnesses, and would eventually die. Relying on an inefficient system of infection, and with the body attacking it from all directions, the chances of viral survival would ordinarily be very slim. Only provirally infected cells that have found a "safe haven" somewhere in the body would have significant longevity. The percentage of viral offspring lucky enough to have survived would be very small. The means Nature commonly uses to overcome such enormous odds against survival is to produce equally enormous numbers of offspring to compensate. This production of huge numbers of progeny results in a huge number of mutations as well, allowing the virus to adapt to its environment quickly and overcome drug treatment. Treatment with medications that block production of GP-120, or that prevent syncytial fusion would prove effective; but only to the extent that they prevent the virus from moving into another cell and producing more provirus. Complete blockage of the syncytial process would halt disease progression. Eventually, provirally infected cells would die, and any GP-160 on their surfaces would degrade to signal the production of new T-cells. If provirally infected cells were long-lived and did NOT readily die off for some reason, the blocking of syncytial fusion would still slow or halt disease progression in spite of elevated viral load counts. However, without the death of these cells, the necessary "replacement signal" would not be sent, and T-cell counts would not rise beyond a certain point. In rare cases, a strain of HIV might exhibit resistance to drug treatment because of the production of a variant of GP-120 that is less efficient. The result would be the otherwise bizarre situation that viral load and T-cell counts might BOTH rise. All of this implies that the level of HIV viral load is a "surrogate" marker, indirectly measuring the number of, and the level of activity of, provirally infected cells. Now, I don't know about you, but that sure SOUNDS like what I've observed first-hand over the years. The THEORY might not be correct in every detail, but it sure has been useful to me as a model for how to think about this disease. At least from a patient's point of view. Three things have caused me to think long and hard before publishing The THEORY:
Okay, # 3 is going to require a little explanation: Howard is this guy who, for years now, has been showing up at medical conferences, vocally advocating the investigation of the use of aspirin in treating HIV infection. Howard claims that significant slowing of the progression of HIV disease can be accomplished by the proper administration of aspirin. He observed this phenomenon himself many years ago. Nobody every seemed to want to listen to Howard. There aren't big bucks to be made off aspirin, so no one wanted to invest money in investigating it. Through a sense of duty to the community, Howard became very vocal about this. He researched the topic, debated doctors and scientists, and stood up at medical conferences to ask for money to do proper clinical trials. Howard has done so much research on aspirin that he is now probably one of the world's leading experts on the topic. His theory about aspirin is so technical I don't even understand it. Thanks to Howard, some limited scientific research on the use of aspirin to treat HIV has been done.The point of the story is this: My initial response upon first hearing Howard bring up the issue at a medical conference was: "Hmmm... another anti-inflammatory." I had been observing for years that anti-inflammatories and immune suppressants such as prednisone and cyclosporin had a positive effect on HIV disease, though no doctor would help investigate them. The response to Howard from the majority of the other participants at that conference was: "Uh-oh...a nut!" Despite years of very good research by Howard, and despite his insistence that aspirin can be used to TREAT HIV disease, not CURE it, Howard is still saddled with the label: "That nut who thinks aspirin cures AIDS." To his credit, Howard continues the fight for aspirin research to this day. The reason Howard is item # 3 above is this: I just didn't want to be labeled as "That nut at Steve's website with that incomprehensible AIDS theory." I have decided, however, that The THEORY presents a pretty comprehensive model of AIDS, and is worth advancing... as a mental exercise, if for no other reason. I guess that makes ME the nutshell that contains The THEORY. So, there you have it. I'm finished. That's my mental exercise for a while. I'm gonna go back to bed and rest up, now. Perhaps, if I get enough rest, I will next attempt to explain why Barney, the Purple Dinosaur is so popular. Naw! I don't think even I could tackle that one! ;-) ...............Dickie
Completed: December, 1998 |